角鼻龙属
角鼻龙属(属名:Ceratosaurus,发音:/ˌsɛrətoʊˈsɔːrəs/,意为“角蜥蜴”)是兽脚类恐龙的一个属,生存年代为晚侏罗世(启莫里阶至提通阶),分布范围包含北美洲、可能还有欧洲、非洲、南美洲等地。角鼻龙是种中型、二足行走的肉食性动物,正模标本估计身长5.3或5.69公尺,凹齿角鼻龙则更大型,可达7公尺长。角鼻龙特色在于很深的颌部,其中生长著比例特长的刀片状牙齿;鼻部中线突出一只脊状角,眼睛上方也有一对角状物;前肢短但功能健全,上有四指;尾部纵向很宽;沿颈部、背部至尾部中线长有一排小型突起的皮甲,并在其馀未知身体部位亦有分布。
| 角鼻龙属 | |
|---|---|

| |
| 克里夫兰罗伊采石场出土的角鼻龙骨架,展示于犹他州自然历史博物馆 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 兽脚亚目 Theropoda |
| 科: | †角鼻龙科 Ceratosauridae |
| 属: | †角鼻龙属 Ceratosaurus Marsh, 1884 |
| 模式种 | |
| †鼻角角鼻龙 Ceratosaurus nasicornis Marsh, 1884
| |
| 其它种 | |
| |
| 异名 | |
| |
1884年美国古生物学家奥斯尼尔·查尔斯·马许根据在科罗拉多花园公园属于莫里逊组地层出土的一副近乎完整的骨骼建立了模式种鼻角角鼻龙(C. nasicornis)。花园公园的标本至今仍是该属已知最完整的骨骼,此后仅有其他少量标本被描述。2000年根据从犹他克里夫兰罗伊采石场与科罗拉多果城找到的两副破碎骨骼描述了另外两个物种:凹齿角鼻龙(C. dentisulcatus)、大角角鼻龙(C. magnicornis),但其有效性备受质疑,这三种的骨骼很可能只是同一物种的不同生长阶段。1999年记载了首个幼体标本发现。2000年描述了葡萄牙洛里尼扬组的部分骨骼,成为角鼻龙在北美以外分布的新证据。坦尚尼亚、乌拉圭、瑞士亦有可能的破碎遗骸曾被提及,但目前多数学者不承认这些属于角鼻龙。
角鼻龙成为兽脚亚目中一大分支角鼻龙类的命名依据,这个演化支在早期即与较接近鸟类的坚尾龙类分化开来,彼此互成姊妹群。在角鼻龙类中,一些学者认为与其最近缘的物种是阿根廷的锐颌龙,后者有著类似的加长牙齿形状。英格兰地质较古老的原角鼻龙曾一度被认为是角鼻龙的直系祖先,现在已知道两者并无关联。
角鼻龙与异特龙、蛮龙等其他侏罗纪北美的代表性大型兽脚类共享栖地,但其数量明显比繁盛的异特龙要罕见许多。有推测这些掠食者的食物偏好彼此有所差异,从而占据不同栖位以减少竞争。角鼻龙可能以植食性恐龙为猎物,捕食鱼类等水生猎物亦曾被提出。鼻角功能可能并非马许最初假设的武器,而作为视觉展示物的可能性更高。
角鼻龙在19世纪末被描述为一种非常致命的代表性肉食恐龙,广泛被当时各大古生物博物馆所青睐,但到了20世纪初,暴龙的发现及媒体文化的加持,使其逐渐取代角鼻龙原本的地位,成为了现代人们对肉食恐龙的代表形象。
发现历史
编辑模式种与正模标本
编辑
首个标本、同时也是正模标本USNM 4735,于1883至1884年由农民费屈(Marshall Parker Felch)所发现并挖掘。[1]发现时状态呈现关节相连、几乎完整并含有头骨。较显著遗失部位包含:未知数量的脊椎、体腔除最后一节之外的所有肋骨、肱骨、双手远端指骨、右前肢大部分、左后肢大部分、脚部大部分。[2]:77骨骼被包覆于坚硬的砂岩之中,头骨与脊椎在化石形成过程中严重扭曲变形。[2]:2,114发现地点位于科罗拉多州峡谷城的花园公园,被称为费屈一号采石场(Felch Quarry 1),属于莫里逊组化石蕴含量最丰富的地层遗址之一。早在角鼻龙发现以前,已在当地挖出大量恐龙化石,其中较著名的包括1883至1884年异特龙近乎完整的正模标本USNM 4734。[2]:7,114
挖掘完后,标本以船运往纽哈芬的皮博迪自然史博物馆,在那里由马许进行研究,1884年发表了新属新种鼻角角鼻龙(Ceratosaurus nasicornis)。[3][2]:114属名Ceratosaurus由古希腊语的κερας/κερατος/keras/keratos(角)加上σαυρος/sauros(蜥蜴);种名nasicornis取自拉丁语的nasus(鼻子)和cornu(角);学名全意即为“鼻子长角的角蜥蜴”。[4]因标本十分完整,发表后角鼻龙很快成为全美最知名的兽脚类恐龙。1898至1899年间,该标本与其他许多由马许描述的标本一同转移至华盛顿特区的美国国立自然史博物馆。抵达当地时,仅少部分材料有清修,随后于1911年起一直持续到1918年底进行了其馀的清修工程。从纽哈芬到华盛顿DC的包装及搬运过程对角鼻龙标本造成了一些程度的损伤。[2]:2,1141920年吉尔摩发表了针对来自纽哈芬的角鼻龙及其他兽脚类标本的全面性重新描述,这之中就包括了同样采石场出土也几乎完整的异特龙。[2]:2

马许在他1892年的论文描绘了角鼻龙首个骨架重建图,身长6.7公尺、身高3.7公尺。[5]吉尔摩在1920年指出,前者的重建把躯干弄得太长,至少多加了六节脊椎。该错误在随后多篇文献中反复提及,包括1899年(1920年才出版)邦德(Frank Bond)在奈特指导下绘制的首件生前复原图。1901年出版更准确的复原图是由格里森同样于奈特监督下制作。吉尔摩于1910至1911年间装设了正模骨架,并展示于国立自然史博馆。多数早期重建都把角鼻龙描绘成尾巴拖地的直立站姿。[2]:115–116相比之下吉尔摩的重建在当时颇为前卫:[6]:276由大腿骨上端得到启发,发现其与小腿成一定角度,而形成水平而非直立、尾部悬空、能够奔跑的姿势。因为化石受压缩变形的程度严重,吉尔摩难以架设立体骨骼,而用浅浮雕的样貌展示于墙面上。[2]:114部分骨骼嵌入墙面,使要进行更深入研究受到阻碍。在2014至2019年博物馆恐龙展示翻修期间,馆方将标本从墙面取下,[7][8]并于2019年计画开幕的新展览中,以一个新制作的立体复制品取而代之;原始标本则回归博物馆馆藏,使学者得以彻底研究。[8]
北美其他发现
编辑自模式种的正模发现以后,直到1960年代才再次发现另一具较重要的角鼻龙化石,是由詹姆斯·麦德森及团队在犹他克里夫兰罗伊采石场挖出破碎、关节脱落的部分骨骼及头骨UMNH VP 5278标本,是体型最大的角鼻龙标本之一。[9]:211976年艾瑞克森(Thor Erikson)在科罗拉多果城发现第二个关节连接、含头骨的标本MWC 1。[10]这是一只相对完整的骨骼,缺乏下颌、手臂及腹肋。虽然头骨较完整,发现时已呈关节脱落状态,侧面压得很扁。来自一只大型个体,但由头骨缝线开放程度显示,其尚未达到成熟的体型。[9]:2–3提供博物馆展示的科学准确3D头骨重建模型,制作过程很复杂,从单一件原始骨骼翻模、矫正变形部位、重建遗失部分、将骨骼模型安装到适当位置、最后上色以还原骨骼原始色彩。[11]
果城和克里夫兰罗伊的标本均于2000年由麦德森和塞缪尔·保罗·威尔斯在著作中进行描述,犹他标本被指定为新种凹齿角鼻龙 (C. dentisulcatus );科罗拉多标本则为大角角鼻龙(C. magnicornis)。[9]前者的种名dentisulcatus指该标本的前上颌骨牙齿及下颌最前面三颗牙齿有平行凹槽;后者的magnicornis则指较大型的鼻角。[9]:2,21这两个物种有效性在后续研究中多次受到质疑。2000年布里特(Brooks Britt)等人表示鼻角种正模标本其实是只少年个体,两个较大型物种则是前者的成年状态。[12]2003年劳赫特(Oliver Rauhut)及2008年卡拉诺(Matthew Carrano)和桑普森(Scott Sampson)评估麦德森和韦尔斯所引用的解剖学特征,来表明这些另外物种只是个体发育(与年龄有关)或个体间形态上的差异。[13][14]:192


1992年在犹他莫尔东南方的玛瑙盆地采石场(Agate Basin Quarry)发现了更进一步的标本BYUVP 12893,目前尚待描述。该标本来自已知最大只的个体。包括头骨前半部、破碎的七节骨盆处脊椎、关节相连的骨盆与荐骨。[14]:192[9]:361999年布里特记载了在怀俄明骨棺采石场发现一只少年角鼻龙个体,由一个完整的头骨、及其馀30%的骨骼部位(包括一个完整的骨盆)组成,其体型小于鼻角种正模的34%。[15]
除了上述五项发现外,还有来自莫里逊组第二、第四至六层带不同地点的零碎角鼻龙遗骸的记载,[16]其中一些位在重要的化石遗址。如犹他恐龙国家纪念区的单独右前上颌骨DNM 972;怀俄明科摩崖的大型肩胛鸟喙骨;科罗拉多旱台采石场的左肩胛鸟喙骨、脊椎和肢骨碎片;科罗拉多米盖特摩尔采石场(Mygatt Moore Quarry)的牙齿。[9]:36
北美以外的发现
编辑非洲
编辑1909至1913年期间,柏林自然博物馆派出德国考察队至德属东非(今坦尚尼亚),从敦达古鲁层挖掘出了多样化恐龙化石。[17]尽管作为非洲最重要的侏㑩纪恐龙化石产地,[17]当地的大型兽脚类恐龙却仅有少量破碎不堪的遗骸得知。[18]1920年德国古生物学家沃纳·亚嫩施将从TL采石场挖到的多节背椎归入角鼻龙未命名种(Ceratosaurus sp.)。1925年亚嫩施根据Mw采石场的破碎遗骸,包括方骨、腓骨、尾椎等碎片,命名了新种罗氏角鼻龙(Ceratosaurus roechlingi),该标本属于一只比鼻角种正模标本比例上更大型的个体。[18]
麦德森和韦尔斯在他们2000年的专著中证实将以上发现归入角鼻龙。此外他们还将原本亚嫩施归入贪食龙可能物种(Labrosaurus (?) stechowi)的牙齿改归入角鼻龙。[9]其他研究人员质疑敦达古鲁层化石的身分,指出这些标本无法辨识出任何角鼻龙的鉴定特征。[19][20]:66[14]:192[21]2011年劳赫特发现罗氏角鼻龙及贪食龙都可能是种角鼻龙科,但缺乏可鉴定为独立属的鉴定特征,因此是疑名。[21]1990年提摩西·罗威(Timothy Rowe)和贾奎斯·高锡尔提到敦达古鲁25颗单独、非常大型(长达15公分)的牙齿,据传曾由亚嫩施提出为另一个物种硕大角鼻龙(Ceratosaurus ingens)。[19][18]然而亚嫩施其实将之归入斑龙而非角鼻龙,也可能是单纯印刷误会。[9]:37[18]2011年劳赫特发现硕大斑龙/角鼻龙实际上与两者都无关连,反而可能是种鲨齿龙科。[21]
欧洲
编辑一个来自英格兰中侏罗世地层的破碎头骨似乎显示出与角鼻龙相似的鼻角。1926年冯休内将头骨描述命名成原角鼻龙,推测是角鼻龙的直系祖先。[22]现在已经知道原角鼻龙其实属于更衍化的暴龙超科;[23]类似的鼻突结构可能是趋同演化。[14]:1852010年劳赫特等人为其建立了专属的原角鼻龙科,且指出保存不完整的“鼻角”实际上只是更大型头冠的最前端断裂,一如近缘五彩冠龙的冠饰结构。[23]
2000年和2006年时,奥塔维欧·马谛尤斯所率领的古生物学团队描述了在葡萄牙中西部洛里尼扬层的化石ML 352,出土自洛里尼扬与托雷斯韦德拉什之间的Valmitão海岸悬崖上,包括一个右股骨、一个左胫骨、以及数个单独的牙齿,作为角鼻龙的一个新种。[24][25]这些骨头埋藏于黄色至棕色细粒径砂岩,为河流泛滥冲积而成,属于新港段(Porto Novo Member)下部层位,地质年代约启莫里阶晚期。该个体的其他骨头SHN (JJS)-65包含左股骨、右胫骨、部分左腓骨,因悬崖持续的侵蚀作用而暴露出来。起初为私人收藏家所有,后来捐赠给托雷斯韦德拉什的自然史协会,使其成为额外的官方标本,并于2015年进行详细描述。[26]2006年马谛尤斯等人将该标本归入凹齿角鼻龙。[25]2008年卡拉诺和桑普森的检验证实的确属于角鼻龙,但尚不足以指定为任何特定物种。[14]:1922015年伊莉莎白·马拉法雅(Elisabete Malafaia)等人质疑凹齿种的有效性,并将标本列在近似鼻角角鼻龙(aff. Ceratosaurus nasicornis)名下。[26]
其他记载像是瑞士穆捷的单一牙齿,起初于1920年由亚嫩施命名为梅氏贪食龙(Labrosaurus meriani),后来麦德森和韦尔斯归入角鼻龙未命名种。[9]:35–362008年马提亚斯·索托(Matías Soto)和丹尼尔·皮瑞(Daniel Perea)描述了乌拉圭塔瓜伦波层的牙齿(推测是前上颌骨牙冠)。其内侧显示出垂直条纹、前缘无锯齿状,这种特征组合仅于角鼻龙得知,然而研究人员指出有鉴于残骸稀缺,将欧洲及非洲材料归入角鼻龙并不可靠,且须对其进一步谨慎评估。[27]2020年索托等人描述了更多来自同地层的角鼻龙牙齿,进而证实他们过去的解释。[28]
特征描述
编辑体型
编辑
角鼻龙符合兽脚类的典型身体结构。[5]角鼻龙是一类手臂缩短、用强有力后腿移动的双足动物。鼻角种首个发现也是正模标本的USNM 4735,不同研究估计出其体长为5.3公尺或5.69公尺。[2]:115[6]该个体是否达成熟则未能确认。[20]:661884年马许推测这个标本体重可能是同期异特龙的一半。[3]较近期的估计则得出418公斤、524公斤、670公斤的数值。[29]后来20世纪下半所发现的另外三只标本体型更大。第一具UMNH VP 5278麦德森曾非正式推估身长达8.8公尺,[10]后来缩减至7公尺。[30]不同研究测量的体重值为980公斤、452公斤、700公斤。[6][30][31]第二具MWC 1略小于前者,体重可能约275公斤。[31]第三具BYUVP 12893尚未经过正式描述,被称是已发现最大型的标本,但数值并未公布。[14]:1922000年葡萄牙发现的ML 352估计身长6公尺及体重600公斤。[30]
头骨
编辑
与身体其馀部分相比,头骨占了相当大的比例。[5]鼻角种正模标本的头骨由吻尖至枕髁(寰椎附著处)测量长55公分。[2]:88该头骨因严重变形使宽度难以准确复原,1920年吉尔摩版本后来被发现复原得过宽。[32]MWC 1近乎完整的头骨测量长60公分及宽16公分;较正模标本来得加长一些。[9]:3头骨孔洞大,使头后部结构比其他大型兽脚类轻,但下颌更深能容纳大比例的牙齿。[6]:277泪骨除了作为眶前孔(眼窝与鼻孔间的大孔)后缘也作为上缘,不同于阿贝力龙科。方骨(底部关节与下颌连接)倾斜,导致颌关节位置相对枕髁更往后移;亦造成侧颞孔(眼窝后方的大孔)底部加宽。[20]:53
角鼻龙最明显的特征在于突出的鼻角,位于骨质鼻孔后方,沿头骨中线处生长,由左右鼻骨愈合构成突起。[2]:82化石仅保存骨质核心部分,相信生前可能覆盖著角质鞘。虽然角核底部平滑,上面三分之二表面布满皱褶与多道凹槽,生前可能分布著血管。正模标本角核长13公分及宽2公分,接著大幅缩减至长7公分及宽1.2公分。[2]:82MWC 1的鼻角则较长且低矮。[9]:3生前若覆盖角质鞘将使鼻角整体更为加长。[33]鼻角后方的鼻骨构成一椭圆凹陷;该凹陷连同鼻角均作为角鼻龙及近缘物种间的区别特征。[14]:192除了大型鼻角,在双眼前方有稍小、半圆形骨质脊状物,由泪骨构成,类似异特龙。[31]年轻个体的三支角都小于成体,且角核两边尚未完全愈合。[15]
_-_AMNH_-_DSC06292.JPG)
构成吻尖的前上颌骨每侧仅三颗牙齿,比许多兽脚类都少。[20]:52正模标本上颌骨两侧各有15颗刀刃状牙齿,前八颗特长且厚实,第九颗起尺寸渐小。如同典型兽脚类,侧缘有小锯齿状,正模标本每5公厘有约10个小锯齿突。[2]:92MWC 1上颌骨仅有11至12颗牙齿、UMNH VP 5278每侧则有12颗;后者牙齿更加厚实与弯曲。[9]:3,27全部标本上颌牙冠都特长;UMNH VP 5278的长达9.3公分,与下颌最矮牙齿同高;正模标本的长7公分而超越下颌最矮齿。其馀兽脚类仅锐颌龙有相似状况。[34]相较之下某些阿贝力龙科牙冠特别短。[20]:92正模标本下颌齿骨每侧有15颗牙齿,保存状况很糟。MWC 1和UMNH VP 5278每侧仅有11颗,后者显示下颌齿相较于上颌比较直且没那么厚实。[9]:3,21
颅后骨骼
编辑
因为正模标本保存的脊柱存在许多段空缺,确切的脊椎数目无法确定。在荐骨之前,颈部及背部由至少20节脊椎构成。颈部中部椎体长且高,前后部椎体则长度短于高度。朝上的神经棘相对大型,背椎神经棘高度与椎体长度相当。荐骨由六个荐椎构成,朝上呈拱状,中部椎体高度大幅缩减,在其他某些角鼻龙类也可见此情况。[20]:55–58尾部占全身长度的一半,由大约50节尾椎构成。正模标本尾巴长2.84公尺。[3][2]:115尾部纵向很高,因为神经棘和人字骨都有加长。如同其他恐龙的尾部功能,作为平衡身体,并含有大型尾股肌,运动时负责向前的推进力,肌肉收缩时将大腿上部往后拉。[20]:55–58

肩胛骨跟鸟喙骨融合成单一块骨头,彼此间无任何明显界线。[20]:58鼻角种正模保存了关节相连的左前肢,含有不完整的手部,虽然清修时造成关节脱落,但研究人员事先制作了石膏模型以记录骨头的原始相对位置。没有任何标本保存下腕骨,使一些研究人员提出角鼻龙可能已经失去这项构造。2016年马修·卡拉诺(Matthew Carrano)和乔纳·休尼尔(Jonah Choiniere)根据模型呈现前臂骨与掌骨之间缝隙及其中的表面纹理来判断,提出存在一或多个软骨型的腕骨的可能性。[35]大多数较进化的兽脚类只有三根手指,但角鼻龙有四根手指,其中第四指明显缩短。第一及第四掌骨短,第二掌骨略长于第三。不同于多数基础兽脚类,掌骨尤其第一指比例特别短。正模标本只保存了第二、三、四指的第一指骨,指骨总数和爪骨未知。第一掌骨结构显示该指生前也存在指骨。脚部由三趾承重,为第二至四趾,通常兽脚类第一趾缩短退化悬空,正模中没有保存下来。1884年马许假设角鼻龙遗失第一趾构造,但1920年吉尔摩指出第二跖骨的附著处证实存在第一趾。[2]:112
角鼻龙在兽脚类中的独特之处在于,沿身体中线生长著小型、加长、形状不规则的皮甲。在第4、5颈椎、第4至10尾椎神经棘上方保存了皮甲,可能构成连续一排,从头骨后方开始延伸至尾巴大部分。1920年吉尔摩推测围岩中呈现的皮甲位置可能反映了生前状态。尾巴上方的皮甲和神经棘间相隔25至38公厘,可能代表其间存在皮肤或肌肉,至于颈部皮甲与神经棘较靠近。除身体中线皮肤还有其他皮甲,如正模的58×70公厘大致四边形骨板,其分布于身体的位置未知。[2]:113–114UMNH VP 5278标本也发现许多皮甲,描述为不规则状。虽然多数皮甲发现位置与骨骼距离不超过5公尺,却与任何脊椎无关连迹象,这和正模的情况不同,难以判断其生前部位。[9]:32
分类系统
编辑
马许在初始论文及后续研究中指出角鼻龙在当时已知兽脚类前所未见的特征。[14]:185其中两项特征:愈合的骨盆、并跖骨,是现代鸟类已知的特征,根据马许所述,这是表明恐龙与鸟类之间关系密切的证据之一。[36]为了将角鼻龙与异特龙、斑龙、虚骨龙区别开来,马许为其建立了新的专属分类群:角鼻龙下目/角鼻龙类与角鼻龙科,并作为底下唯一物种。[14]:1851892年马许在化石战争的死对头寇普对其观点提出质疑,他认为鼻角等独有特征仅能支持作为一个新种,而无法区别成为独立的属,于是他将鼻角种归入斑龙属,成为鼻角斑龙(Megalosaurus nasicornis)。[37]

虽然后续研究皆视角鼻龙为一独立属,[2]:76在接下来一个世纪以来其亲缘关系持续引发争议。角鼻龙科和角鼻龙下目都未被广泛接受,且仅有其他少数几种所知甚少的物种被发现。多年来角鼻龙曾在不同文献被归入各种分类,如恐齿龙科、斑龙科、虚骨龙类、肉食龙下目、恐齿龙超科。[9]:21920年吉尔摩的重新检验认为角鼻龙是三叠纪之后已知最原始的兽脚类,与当时已知任何其他同时代的兽脚类都不近缘,理应分在自己专属的角鼻龙科中。[2]:76但直到1980年代系统发生学分析的建立,马许将角鼻龙类作为一独特分类群的观点才终于得到支持。1985年南美新发现的阿贝力龙和食肉牛龙被发现与角鼻龙近缘。1986年高锡尔辨识出腔骨龙超科与角鼻龙近缘,不过在近期研究该演化支通常被排除于角鼻龙类之外。自此之后,更多角鼻龙类的新物种相继被发现。[14]:185

角鼻龙类从兽脚类演化阶段的非常早期就与另一条演化至鸟类的坚尾龙类分化开来,于是常被当作相对基础的演化支。[38]角鼻龙类的范畴底下包含了两个更进化的分支西北阿根廷龙科和阿贝力龙科,两者一同位于阿贝力龙超科内;另外还有一些较基础的物种,如轻龙、三角洲奔龙、角鼻龙。角鼻龙本身在基础角鼻龙类的确切位置至今仍有争议。某些分析认为角鼻龙是所有基础物种中相对最进阶的,并与阿贝力龙超科互成姊妹群。[14]:187[39]2004年奥利弗·劳赫特(Oliver Rauhut)提出锐颌龙与角鼻龙构成姊妹关系,因为两属都以特长的上颌齿为鉴定特征,他把两属归到角鼻龙科,[34]后续研究皆采纳此观点,[40][41][42][33]如以下演化树取自2012年迭哥波和劳赫特:[40]
| 角鼻龙类 Ceratosauria |
| |||||||||||||||||||||||||||||||||||||||||||||
2017年王硕等人认为西北阿根廷龙科并非如过去所想的处于阿贝力龙超科底下,而是比角鼻龙更为基础。因为西北阿根廷龙科已被当作定义阿贝力龙超科及阿贝力龙科的固定依据,于是这样的调整将使得每个演化支容纳进更大范围的分类群,其中也包含角鼻龙本身。2018年拉斐尔·德寇特(Rafael Delcourt)的后续研究支持前述论点,但指出需用从来都很少使用的角鼻龙超科来当作阿贝力龙超科的异名取代之。至于阿贝力龙科,德寇特为其指定出一种排除掉角鼻龙的新定义,而得以延用旧名。王等人还另外发现角鼻龙、锐颌龙、阿根廷的始阿贝力龙构成一个演化支。[42]德寇特以角鼻龙科代指这个范围的演化支,并建议将角鼻龙科的定义修正为与角鼻龙接近、而与食肉牛龙较远的所有物种。其支序图如下所示:[33]
| 角鼻龙超科 Ceratosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物学
编辑栖位与食性
编辑
在莫里逊层与洛里尼扬层,角鼻龙的化石经常与其他大型兽脚类一同发现,包括斑龙科的蛮龙、[43]异特龙科的异特龙。科罗拉多花园公园除了角鼻龙也有异特龙。科罗拉多旱台采石场及犹他克里夫兰罗伊采石场分别发现至少三种大型兽脚类:角鼻龙、异特龙、蛮龙。[32][44]同样在怀俄明科摩崖及邻近地区也产出角鼻龙、异特龙、还有至少一种大型斑龙科。[45]角鼻龙是这些兽脚类动物群中数量极其稀有者,在异特龙和角鼻龙分布重叠区域,前者数量几乎是后者的7.5倍之多。[46]
许多篇研究尝试如何解释这些共栖掠食者如何降低直接竞争机率。1998年唐纳·韩德森(Donald Henderson)提到角鼻龙和至少两个潜在的异特龙物种共存,称后两者在形态上有所分化:一种吻部缩短、头骨较高且宽、牙齿短而后弯;另一种吻部较长、头骨低矮、牙齿长而笔直。总体来说,若共栖物种之间的形态、生理构造、生活行为等方面的相似程度越高,彼此竞争就越激烈。韩德森结论道:短吻异特龙与长吻异特龙、角鼻龙有著不同栖位;短吻形态减少咬合产生的应力从而增加咬合力,类似猫科的状况。至于长吻型异特龙和角鼻龙的长窄吻部及长尖牙则适合迅速猛烈的小范围咬合,类似犬科。韩德森指出由于角鼻龙和长吻异特龙的头骨形态十分相似,显示两者可能会直接竞争。根据角鼻龙在克里夫兰罗伊采石场的罕见程度,以及当地长吻异特龙的繁盛情况来判断,前者可能受不了后者竞争而几乎被赶出栖地。但花园公园和旱台的角鼻龙似乎比较常见,并与短吻异特龙共存。[32]

另外韩德森认为角鼻龙也许能够透过不一样的猎物偏好来避免竞争;其演化出极细长的牙齿可能是与长吻异特龙竞争下的产物,在作为食腐者时两者可能选择取食尸体的不同部位。角鼻龙的加长牙齿也能当作物种间的视觉识别或其他社会功能。再者这些兽脚类同样趋向体型巨大化也会降低竞争机率,因为当体型增加能选择的猎物种类、范围亦随之增广。[32]
2006年福斯特和丹尼尔·舒尔(Daniel Chure)认同韩德森的理论:有鉴于角鼻龙和异特龙共栖并捕食同类型猎物的事实,所以可能各自采取不同社食策略以避免竞争。据他们表示头骨、牙齿、前肢在比例上有所差异也代表了证据支持。[46]但后续研究则质疑了异特龙的两种变异形态差异。2010年肯尼思·卡彭特发现克里夫兰罗伊采石场的短吻异特龙反映了极端程度的个体差异,而非不同物种的变异。[47]而且韩德森提出短吻形态的依据-花园公园的USNM 4734头骨标本,实际上是将样貌复原得太短的误会。[48]
2004年罗伯特·巴克和盖瑞·比尔(Gary Bir)提出角鼻龙主要特化来觅食水生猎物,如肺鱼、鳄鱼、海龟。统计了科摩崖及周围50个地点发现的脱落牙齿,显示角鼻龙和斑龙科的牙齿在水域邻近区域(潮湿的泛滥平原、湖岸、沼泽)最常见。角鼻龙偶尔出现于内陆环境。至于异特龙科无论是内陆或近水都相当普遍。巴克和比尔据此结论道:角鼻龙和斑龙科必定在水域附近环境捕猎,而角鼻龙偶尔也会去内陆吃大型恐龙尸体。研究者还注意到角鼻龙与斑龙科都有著狭长、低矮、灵活的身体。与其他莫里逊层兽脚类相比,角鼻龙前部尾椎神经棘高耸、垂直而非后倾,加上尾椎下方的长型人字骨,显示可能构成一条如鳄鱼般的纵深形尾部,适合游泳。异特龙则相反,身形短高、粗壮结实、腿长,适应于开阔地区奔驰并追捕大型植食恐龙(如蜥脚类和剑龙类),但巴克和比尔推测在这些猎物随季节发生短缺时,异特龙也会投机转而觅食水生猎物。[45]但该理论受到2019年尹灿奎的质疑,他表示角鼻龙仅比莫里逊层其他兽脚类更有能力捕捉水生猎物,而非完全半水栖生活。[49]
1986年巴克的畅销书《恐龙异说》(The Dinosaur Heresies)中认为上颌骨仅松散附著于周围的头骨上,能够在颅内产生一定程度的活动范围,称为头骨运动。同样地,下颌可以相互移动,方骨外扩,使下颌于关节处撑开,让动物能张大嘴巴以吞下大型食物。[50]2008年凯西·郝立蝶(Casey Holliday)和罗伦斯·灰墨(Lawrence Witmer)重新审视这项关于其他恐龙的假说,认为没有任何证据支持恐龙存在肌肉驱动的头骨运动,甚至多数恐龙都没办法做到。[51]

在一只异特龙的耻骨脚(耻骨末端扩张处)发现某种别的兽脚类齿痕,可能由角鼻龙或蛮龙造成。因为该骨头位于自身难以构到的身体底缘并是整个身体最大块骨头,代表是这只异特龙尸体被啃食。[52]麦杰特摩尔(Mygatt-Moore)采石场保存了异常高频率的兽脚类咬痕,其中大部分来自异特龙和角鼻龙,也有更大的蛮龙或食蜥王龙。虽然植食动物尸体上的咬痕位置与被杀时或刚开始吃的位置一致,但异特龙身上也找到了其他异特龙或兽脚类留下的啃尸痕迹。与其他化石群相比之下,当地异常高密度的兽脚类咬痕,可能代表著旱季来临掠食者们争相抢夺紧缩的食物来源,或仅归咎于其他化石点的采样偏差。[53]
鼻角与皮甲功能
编辑1884年马许认为角鼻龙的鼻角是作为攻击与防御的强大武器,1920年吉尔摩认同其论点。[3]:331[2]:82然而现代普遍认为鼻角不太可能当作武器。[10]1985年大卫·诺曼认为鼻角可能并非用来抵御其他掠食者,而是争夺配偶的雄性角鼻龙种内打斗使用。[54]1988年奎格里·保罗提出类似功能,并绘制两只角鼻龙进行一场非致命的互撞竞赛。[6]1990年罗威和高席尔提出更进一步的论点,认为鼻角仅具有视觉展示功能,完全无法打斗。[19]为了达到炫耀目的,鼻角可能具有鲜艳色彩。[31]另外沿身体中线生长的成排皮甲也可能有展示功能。[19]
前肢功能
编辑
角鼻龙掌骨及指骨大幅缩短,产生疑问关于手部是否仍具其他基础兽脚类推测的抓握功能。角鼻龙类的阿贝力龙科、泥潭龙可观察到更极端的前肢萎缩,完全失去原来的手部功能。[55]2016年卡拉诺和乔纳·休尼尔(Jonah Choiniere)的角鼻龙手部解剖学研究中,指出其仍与其他基础兽脚类形态大致雷同,就算缩短了仍保有基础的抓握能力。虽然只有第一指骨保存下来,但根据上面的发达关节面所示,第二指骨可以灵活移动,甚至可能手指也能做到像其他基础兽脚类那样程度的运动。如同其他兽脚类(不含阿贝力龙科),第一指弯曲时会稍微内旋。[35]
脑部与感官
编辑
马许可能在清修正模化石的过程中监督了脑腔模型的制作,从而得出脑部尺寸中等,但相对植食动物的要大得多。但后来头骨被黏合在一块,使后续研究无法检验该模型的准确程度。[3][2]:93
果城的MWC 1标本保存了第二件较良好的脑壳,由山德斯(Kent Sanders)和史密斯(David Smith)进行CT扫描以重建内耳结构。2005年研究人员表示角鼻龙有著一颗典型的基础兽脚类脑腔,近似于异特龙。参与嗅觉的嗅球印痕保存良好,虽然类似异特龙,但小于暴龙;后者认为有相当灵敏的嗅觉。参与平衡感的半规管能藉以推测习惯性的头部姿态,类似其他兽脚类;通常兽脚类中的情况雷同,显示不同物种彼此有著差不多的运动功能。半规管最前方扩张,常见于二足动物。半规管外侧的方向代表自然状态下头颈维持水平向。[56]
鼻角种正模的左边第二至第四跖骨愈合。1884年马许写了一篇短文探讨这项当时在恐龙前所未见的特征,并提到跟现代鸟类非常相似。[36]1890年关于角鼻龙该特征的存在引发争议,乔格·鲍尔推测这是骨折愈合造成的。1892年寇普认同前述论点,还认为各属间缺乏解剖学差异,应将鼻角种视为斑龙的一种。[37]但自此后已在其他恐龙(包括比角鼻龙更基础的物种)也观察到非病理因素的并跖骨。[57]1920年奥斯本说明就算并跖骨不寻常,却没发现任何明显的骨骼异常发育,可能与病理无关。[2]:1121976年拉克维奇(Ronald Ratkevich)表示跖骨愈合将限制动物奔跑能力,1988年保罗则反驳,指出现今许多移动迅速的动物(如地栖鸟类和有蹄类)也存在并跖骨。[6]1999年坦克(Darren Tanke)和罗斯柴尔德(Bruce Rothschild)进行分析发现该并跖骨的确为病理因素,证实早先鲍尔的论点。[57]其他与角鼻龙有关的病理报告还有脚部的疲劳性骨折、[58]未知种的一颗断牙呈现持续磨耗迹象。[57]
古生态学与古地理学
编辑年代与环境
编辑
所有北美的角鼻龙都出土于莫里逊层,这是美西一系列由浅海和冲积层组成的沉积岩,也是大陆上恐龙化石最繁盛的产地。根据放射性定年法测量该岩层年代范围,从最底层的1亿5630万年前[59]到最顶层的1亿4680万年前[60],横跨晚牛津阶、启莫里阶、到早提通阶。已知的角鼻龙来自启莫里阶和提通阶层带。[20]:49莫里逊层的环境为干湿季分明的半干旱气候。莫里逊盆地一路由新墨西哥州延伸到亚伯达省和萨克齐万省,是洛矶山脉前山开始西推时形成。东向流域盆地的沉积物由溪流、河川所搬运,沉积成低地沼泽、湖泊、河道、泛滥平原。[61]莫里逊层的年代与葡萄牙洛里尼扬层、坦尚尼亚敦达古鲁层大致相近。[62]
北美
编辑莫里逊层保存了由巨型蜥脚类占据优势的环境与时代记录。[63]与角鼻龙共存的恐龙包括:兽脚类剖齿龙、史托龙、长臂猎龙、虚骨龙、马许龙、嗜鸟龙、异特龙、食蜥王龙、蛮龙;蜥脚类迷惑龙、雷龙、春雷龙、载域龙、简棘龙、腕龙、圆顶龙、梁龙、小梁龙、重龙、超龙、极巨龙;装甲类剑龙、西龙、阿尔科瓦龙、怪嘴龙、迈摩尔甲龙;鸟脚类侏儒龙、橡树龙、弯龙。[64]异特龙标本数量占莫里逊层所有兽脚类的70至75%,处于当地食物链顶端。[44]其他共享栖地的脊椎动物还有辐鳍鱼、青蛙、蝾螈、乌龟的多塞龟、喙头目、蜥蜴、陆生和水生鳄形超目如跳鳄、数种翼龙类包括抓颌龙和买萨翼龙。双壳纲、水生蜗牛的壳也很常见。藻类、真菌、藓苔、木贼、苏铁、银杏、数种针叶树化石透露了当时的植相状况。植被景观包括树蕨等蕨类构成的滨岸林及充斥著南洋杉属之类的稀树莽原。[65]
欧洲
编辑葡萄牙洛里尼扬层新港段(Porto Novo Member)发现角鼻龙的破碎遗骸透露了该物种也许存在于此地。洛里尼扬层的恐龙生态组成结构与莫里逊层几乎一样,或至少都是类似物种。[62]除了角鼻龙,葡萄牙岩层也同样出土过来自北美的异特龙和蛮龙,至于洛里尼扬诺龙只在葡萄牙发现过。新港段的植食性恐龙包括蜥脚类的迪涅鲁龙、洛里尼扬龙、兹必龙;剑龙类的米拉加亚龙。[66][24][25]在晚侏罗世,欧洲与北美之间仅隔著刚形成的狭窄大西洋,而葡萄牙所在的伊比利半岛当时是一座大型孤岛,与欧洲其他地区隔离开来。根据马谛尤斯等人所述,葡萄牙与北美兽脚类种群的相似程度,代表之间曾经存在过陆桥使动物得以互相迁徙。[24][25]不过马拉法雅等人提出不同、更为复杂的情境假设,因为葡萄牙其他动物如蜥脚类、海龟、鳄类都与北美有明显差异,代表当陆块开始分开时,某些物种是透过交换迁徙来的,某些则在当地产生特有化。[26]
文化意义
编辑公众形象
编辑角鼻龙是知名的恐龙之一。因为来自美国本土,又早在19世纪就发表,足以长期发展成为科普媒体中代表性的经典史前物种。史密松宁学会博物馆的展览品提供了加持,因为当时那边的恐龙厅展品很少,而角鼻龙独特的鼻角和背脊赋予了一种迷人的“龙状”外表令人印象深刻。[67]
绘画
编辑.jpg)


1899年邦德(Frank Bond)在威柏·克林顿·奈特指导下绘制了可能是历史上首个角鼻龙复原图,但直到1920年才透过吉尔摩公开。[2]1901年费德里克·奥古斯特斯·卢卡斯出版一本关于灭绝动物的科普书《Animals of the Past》,当中有著名童书画乔瑟夫·格里森的复原图,[68]他在查尔斯·R·奈特监督下绘制。吉尔摩曾表示这是最为成功的复原,还将图放在博物馆骨架旁展示。[2]1911年亨利·内维尔·哈钦森的《Extinct Monsters and Creatures of Other Days: a popular account of some of the larger forms of ancient animal life》出现由乔瑟夫·斯密绘的角鼻龙插画,显然参考了马许的错误复原。[69]1915年吉尔摩制作了角鼻龙的黏土模型。[70]
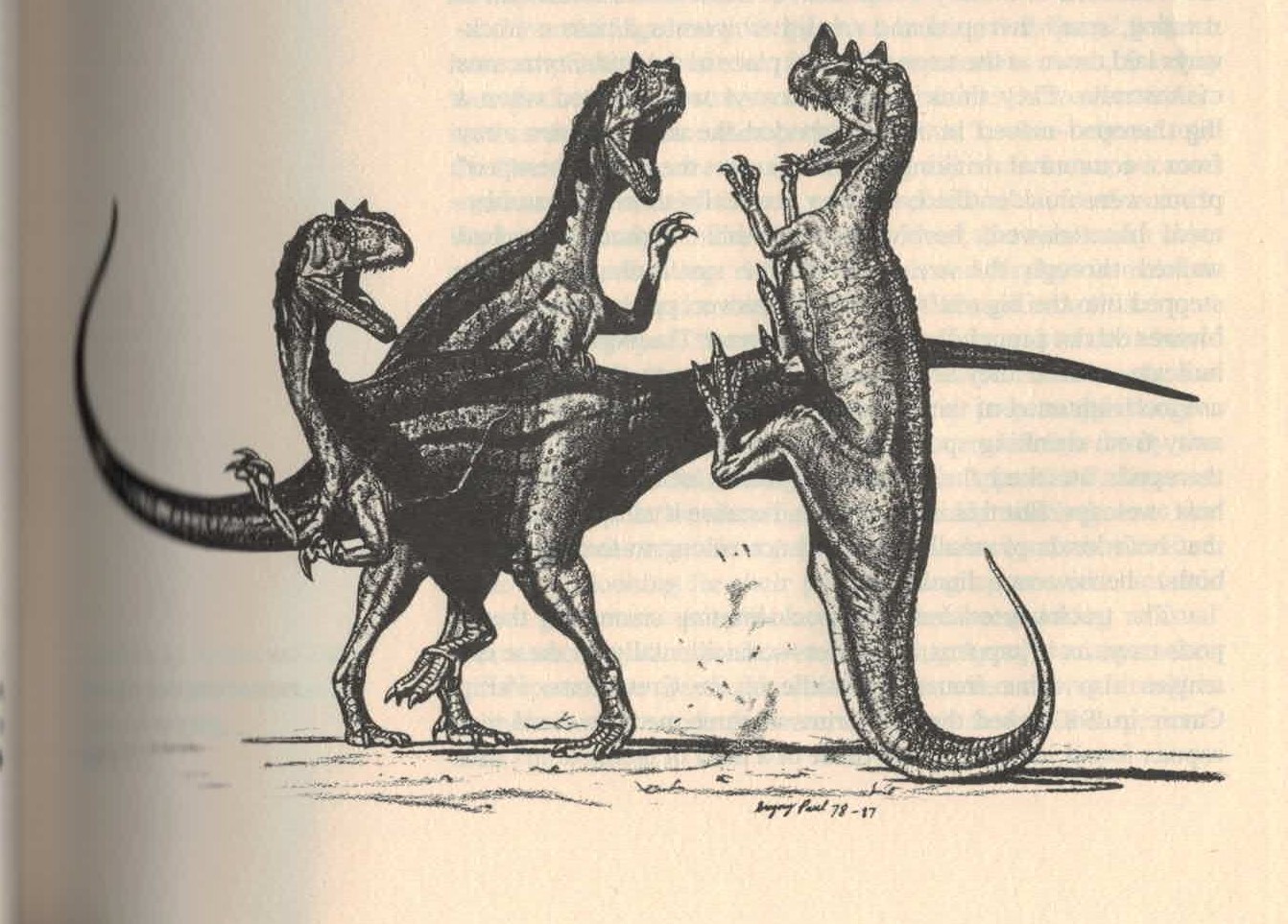
1920年代以后,角鼻龙形象长期固化,后续的画家皆参考格里森或斯密,逐渐完全脱离动物的真实形象。直到1980年代突然起了变革,奎格里·保罗观察了正模标本,将角鼻龙诠释为行动活跃的水平姿态,更贴近现代形象。其中一幅 (页面存档备份,存于互联网档案馆)角鼻龙以尾巴撑地抬起双脚踢击两只异特龙,类似袋鼠的行为。另一幅 (页面存档备份,存于互联网档案馆)依照他对鼻角功能的论点,展现了角鼻龙透过鼻角侧击对抗同类。这些成为新式古生物形象的代表性作品,并在1990年代占据主流地位。[6]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
媒体作品
编辑
角鼻龙早期在文学作品的亮相,可追溯到1918年爱德加·莱斯·巴勒斯的科幻小说《时光遗忘之境》(The Land That Time Forgot)。[71]1942年迪诺·布扎蒂所创作的美文中篇小说《屠龙记》(l'Uccisione del drago)中描述了最后幸存的角鼻龙。[72]角鼻龙亦成为某些神秘生物学的题材,在1908年乔吉斯·德派的作品《鹧鸪溪怪兽》(The Monster of Partridge Creek)描绘育空地区出没的大型二足怪物形象即参考了角鼻龙;[73][74]但后世批判了恐龙生活在极圈的可能性。[75]
20世纪角鼻龙开始经常出现于大萤幕,使肉食恐龙的代表形象广泛为人所知。1914年D·W·格里菲斯的电影《原始人》(Brute Force)中角鼻龙作为威胁穴居部落的史前动物之一,该作因使用机械模型而被视为史上第一部有实体恐龙登场的电影。[76]1940年迪士尼的《幻想曲》(Fantasia)中的《春之祭》片段 (页面存档备份,存于互联网档案馆),角鼻龙攻击陷入泥淖的蜥脚类和剑龙。1948年的电影《未知岛 (页面存档备份,存于互联网档案馆)》也出现角鼻龙。1966年的电影《史前百万年 (页面存档备份,存于互联网档案馆)》有角鼻龙跟三角龙的打斗场景,但这两种实际上不可能共存;片中的恐龙由雷·哈利豪森的定格动画所制作。1975年改编同名小说的电影《时光遗忘之境》《The Land That Time Forgot》及1977年的续集《时光遗忘之人》也出现了角鼻龙。2001年的《侏罗纪公园3》(Jurassic Park III)一只红色的角鼻龙于主角们在棘龙粪便寻找电话的片段中短暂登场。同系列动画《白垩纪冒险营》(Camp Cretaceous)角鼻龙再度回归。[77]角鼻龙亦在几部科普作品有亮相。1956年的纪录片《动物世界》(The Animal World)以定格动画展现角鼻龙杀死剑龙,后同类争食双双跌下悬崖被冲走。[78]2001年探索频道的纪录片《恐龙纪元》(When Dinosaurs Roamed America)的晚侏罗世段落,角鼻龙先登场追捕橡树龙并杀死一只,然后试图袭击剑龙被打倒而放弃,最后再度追捕橡树龙时被异特龙偷袭咬死。[79]
参考来源
编辑- ^ Brinkman, P.D. The Second Jurassic Dinosaur Rush. Museums and Paleontology in America at the Turn of the Twentieth Century. University of Chicago Press. 2010: 10. ISBN 978-0-226-07472-6.
- ^ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 2.11 2.12 2.13 2.14 2.15 2.16 2.17 2.18 2.19 2.20 2.21 2.22 Gilmore, C.W. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus (PDF). Bulletin of the United States National Museum. 1920, 110 (110): 1–154 [2022-06-20]. doi:10.5479/si.03629236.110.i. hdl:2027/uiug.30112032536010
 . (原始内容存档 (PDF)于2021-11-09).
. (原始内容存档 (PDF)于2021-11-09).
- ^ 3.0 3.1 3.2 3.3 3.4 Marsh, O.C. Principal characters of American Jurassic dinosaurs, part VIII: The order Theropoda (PDF). American Journal of Science. 1884, 27 (160): 329–340 [2022-06-20]. Bibcode:1884AmJS...27..329M. doi:10.2475/ajs.s3-27.160.329. (原始内容存档 (PDF)于2023-02-15).
- ^ Simpson, D.P. Cassell's Latin Dictionary 5. London: Cassell Ltd. 1979: 153, 387 [1854]. ISBN 978-0-304-52257-6.
- ^ 5.0 5.1 5.2 Marsh, O.C. Restorations of Claosaurus and Ceratosaurus. American Journal of Science. 1892, 44 (262): 343–349 [2022-06-20]. Bibcode:1892AmJS...44..343M. doi:10.2475/ajs.s3-44.262.343. hdl:2027/hvd.32044107356040 . (原始内容存档于2018-07-22).
- ^ 6.0 6.1 6.2 6.3 6.4 6.5 6.6 Paul, Gregory S. Ceratosaurs. Predatory Dinosaurs of the World. Simon & Schuster. 1988: 274–279. ISBN 978-0-671-61946-6.
- ^ Jacqueline, T. David Koch Donates $35 Million to National Museum of Natural History for Dinosaur Hall. The Washington Post. May 3, 2012 [December 4, 2017]. (原始内容
 存档于2017-07-19).
存档于2017-07-19).
- ^ 8.0 8.1 The Good, Bad and Ugly: Dismantling Historic Fossil Displays, Part Two. Digging the Fossil Record: Paleobiology at the Smithsonian. Smithsonian National Museum of Natural History. [December 4, 2017]. (原始内容存档于October 10, 2017).
- ^ 9.00 9.01 9.02 9.03 9.04 9.05 9.06 9.07 9.08 9.09 9.10 9.11 9.12 9.13 9.14 Madsen, J.H.; Welles, S.P. Ceratosaurus (Dinosauria, Theropoda): A Revised Osteology. Utah Geological Survey. 2000: 1–80. ISBN 978-1-55791-380-7.
- ^ 10.0 10.1 10.2 Glut, D.F. Ceratosaurus. Dinosaurs: The Encyclopedia. McFarland & Company. 1997: 266–270. ISBN 978-0-89950-917-4.
- ^ Tidwell, V. Restoring crushed Jurassic dinosaur skulls for display. The Continental Jurassic. Museum of Northern Arizona Bulletin. 1996, 60.
- ^ Britt, B.B.; Chure, D.J.; Holtz, T.R., Jr.; Miles, C.A.; Stadtman, K.L. A reanalysis of the phylogenetic affinities of Ceratosaurus (Theropoda, Dinosauria) based on new specimens from Utah, Colorado, and Wyoming. Journal of Vertebrate Paleontology. 2000, 20 (suppl): 32A. doi:10.1080/02724634.2000.10010765.
- ^ Rauhut, O.W.M. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology. 2003: 25.
- ^ 14.00 14.01 14.02 14.03 14.04 14.05 14.06 14.07 14.08 14.09 14.10 Carrano, M.T.; Sampson, S.D. The Phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 2008, 6 (2): 183–236. ISSN 1477-2019. doi:10.1017/S1477201907002246.
- ^ 15.0 15.1 Britt, B.B.; Miles, C.A.; Cloward, K.C.; Madsen, J.H. A juvenile Ceratosaurus (Theropoda, Dinosauria) from Bone Cabin Quarry West (Upper Jurassic, Morrison Formation), Wyoming. Journal of Vertebrate Paleontology. 1999, 19 (Supplement to No 3): 33A. doi:10.1080/02724634.1999.10011202.
- ^ Foster, J. Appendix. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 327–329. ISBN 978-0-253-34870-8.
- ^ 17.0 17.1 Zils, C.W.; Moritz, A.; Saanane, C. Tendaguru, the most famous dinosaur locality of Africa. Review, survey and future prospects. Documenta Naturae. 1995, 97: 1–41.
- ^ 18.0 18.1 18.2 18.3 Janensch, W. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica. 1925,. Supplement VIII: 1–100 (德语).
- ^ 19.0 19.1 19.2 19.3 Rowe, T.; Gauthier, J. Ceratosauria. Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). The Dinosauria. University of California Press. 1990: 151–168. ISBN 978-0-520-06726-4.
- ^ 20.0 20.1 20.2 20.3 20.4 20.5 20.6 20.7 20.8 Tykoski, R.S.; Rowe, T. Ceratosauria. Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). The Dinosauria: Second Edition. University of California Press. 2004: 47–70. ISBN 978-0-520-24209-8.
- ^ 21.0 21.1 21.2 Rauhut, O.W.M. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Special Papers in Palaeontology. 2011, 86: 195–239.
- ^ Huene, F.v. On several known and unknown reptiles of the order Saurischia from England and France. Annals and Magazine of Natural History. 1926,. Serie 9 (17): 473–489. doi:10.1080/00222932608633437.
- ^ 23.0 23.1 Rauhut, O.W.M.; Milner, A.C.; Moore-Fay, S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological Journal of the Linnean Society. 2010, 158 (1): 155–195. doi:10.1111/j.1096-3642.2009.00591.x .
- ^ 24.0 24.1 24.2 Mateus, O.; Antunes, M.T. Ceratosaurus (Dinosauria: Theropoda) in the Late Jurassic of Portugal. 31st International Geological Congress, Abstract Volume (Rio de Janeiro). 2000.
- ^ 25.0 25.1 25.2 25.3 Mateus, O.; Walen, A.; Antunes, M.T. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science, Bulletin 36. 2006.
- ^ 26.0 26.1 26.2 Malafaia, E.; Ortega, F.; Escaso, F.; Silva, B. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology. October 3, 2015, 27 (7): 938–946. ISSN 0891-2963. doi:10.1080/08912963.2014.915820.
- ^ Soto, M.; Perea, D. A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Uruguay. Journal of Vertebrate Paleontology. 2008, 28 (2): 439–444. doi:10.1671/0272-4634(2008)28[439:acdtft]2.0.co;2.
- ^ Soto, Matías; Toriño, Pablo; Perea, Daniel. Ceratosaurus (Theropoda, Ceratosauria) teeth from the Tacuarembó Formation (Late Jurassic, Uruguay). Journal of South American Earth Sciences. November 1, 2020, 103: 102781. Bibcode:2020JSAES.10302781S. ISSN 0895-9811. S2CID 224842133. doi:10.1016/j.jsames.2020.102781 (英语).
- ^ Therrien, F.; Henderson, D.M. My theropod is bigger than yours ... or not: estimating body size from skull length in theropods. Journal of Vertebrate Paleontology. 2007, 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:mtibty]2.0.co;2.
- ^ 30.0 30.1 30.2 Paul, Gregory S. The Princeton Field Guide to Dinosaurs 2nd. Princeton University Press. 2016: 84. ISBN 978-0-691-16766-4.
- ^ 31.0 31.1 31.2 31.3 Foster, J. Gargantuan to Minuscule: The Morrison Menagerie, Part II. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 162–242. ISBN 978-0-253-34870-8.
- ^ 32.0 32.1 32.2 32.3 Henderson, D.M. Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods. Gaia. 1998, (15): 219–226.
- ^ 33.0 33.1 33.2 Delcourt, Rafael. Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific Reports. 2018, 8 (1): 9730. Bibcode:2018NatSR...8.9730D. PMC 6021374 . PMID 29950661. doi:10.1038/s41598-018-28154-x.
- ^ 34.0 34.1 Rauhut, O.W.M. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. Journal of Vertebrate Paleontology. 2004, 24 (4): 894–902. doi:10.1671/0272-4634(2004)024[0894:paaogs]2.0.co;2.
- ^ 35.0 35.1 Carrano, M.T.; Choiniere, J. New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution. Journal of Vertebrate Paleontology. 2016, 36 (2): –1054497. doi:10.1080/02724634.2015.1054497.
- ^ 36.0 36.1 Marsh, O.C. On the united metatarsal bones of Ceratosaurus. American Journal of Science. 1884, 28 (164): 161–162 [2022-06-20]. Bibcode:1884AmJS...28..161M. doi:10.2475/ajs.s3-28.164.161. (原始内容存档于2020-06-24).
- ^ 37.0 37.1 Cope, E.D. On the Skull of the Dinosaurian Lælaps incrassatus Cope. Proceedings of the American Philosophical Society. 1892, 30 (138): 240–245. ISSN 0003-049X. JSTOR 983173.
- ^ Hendrickx, C.; Hartman, S.A.; Mateus, O. An overview of non-avian theropod discoveries and classification. PalArch's Journal of Vertebrate Palaeontology. 2015, 12 (1): 1–73.
- ^ Xu, X.; Clark, J.M.; Mo, J.; Choiniere, J.; Forster, C.A.; Erickson, G.M.; Hone, D.W.; Sullivan, C.; Eberth, D.A.; Nesbitt, S.; Zhao, Q. A Jurassic ceratosaur from China helps clarify avian digital homologies (PDF). Nature. 2009, 459 (7249): 940–944 [2022-06-20]. Bibcode:2009Natur.459..940X. PMID 19536256. doi:10.1038/nature08124. (原始内容存档 (PDF)于2017-09-22).
- ^ 40.0 40.1 Pol, D.; Rauhut, O.W.M. A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences. 2012, 279 (1804): 3170–5. PMC 3385738 . PMID 22628475. doi:10.1098/rspb.2012.0660.
- ^ Delcourt, R. Revised morphology of Pycnonemosaurus nevesi Kellner & Campos, 2002 (Theropoda: Abelisauridae) and its phylogenetic relationships. Zootaxa. 2017, 4276 (1): 1–45. PMID 28610214. doi:10.11646/zootaxa.4276.1.1.
- ^ 42.0 42.1 Wang, S.; Stiegler, J.; Amiot, R.; Wang, X.; Du, G.-H.; Clark, J.M.; Xu, X. Extreme ontogenetic changes in a ceratosaurian theropod (PDF). Current Biology. 2017, 27 (1): 144–148. PMID 28017609. doi:10.1016/j.cub.2016.10.043 .
- ^ Hendrickx, Christophe; Mateus, Octávio. Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods. PLOS ONE. March 5, 2014, 9 (3): e88905. Bibcode:2014PLoSO...988905H. ISSN 1932-6203. PMC 3943790 . PMID 24598585. doi:10.1371/journal.pone.0088905 (英语).
- ^ 44.0 44.1 Foster, J.R. Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.. New Mexico Museum of Natural History and Science Bulletin 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2003: 29.
- ^ 45.0 45.1 Bakker, Robert T.; Bir, Gary. Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness. Currie, P.J.; Koppelhus, E.B.; Shugar, M.A.; Wright, J.L. (编). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington and Indianapolis: Indiana University Press. 2004: 301–342. ISBN 978-0-253-34373-4.
- ^ 46.0 46.1 Foster, J.R.; Chure, D.J. Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution. New Mexico Museum of Natural History and Science Bulletin. 2006, 36: 119–122.
- ^ Carpenter, Kenneth. Variation in a population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA. Paleontological Research. 2010, 14 (4): 250–259. doi:10.2517/1342-8144-14.4.250.
- ^ Carpenter, Kenneth; Paul, Gregory S. Comment on Allosaurus Marsh, 1877 (Dinosauria, Theropoda): proposed conservation of usage by designation of a neotype for its type species Allosaurus fragilis Marsh, 1877. Bulletin of Zoological Nomenclature. 2015, 72: 1.
- ^ Changyu Yun. Comments on the ecology of Jurassic theropod dinosaur Ceratosaurus (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle (PDF). Volumina Jurassica. 2019, XVII: 111–116 [2022-07-02]. (原始内容 (PDF)存档于2022-07-12).
- ^ Bakker, R.T. The Dinosaur Heresies 2. William Morrow and Company. 1986: 523. Bibcode:1987Palai...2..523G. ISBN 978-0-688-04287-5. JSTOR 3514623. doi:10.2307/3514623.
|journal=被忽略 (帮助);|issue=被忽略 (帮助) - ^ Holliday, C.M.; Witmer, L.M. Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology. December 12, 2008, 28 (4): 1073–1088. doi:10.1671/0272-4634-28.4.1073.
- ^ Chure, Daniel J.; Fiorillo, Anthony R.; Jacobsen, Aase. Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic. Gaia. 1998, 15: 227–232 [2022-07-02]. (原始内容存档于2021-11-09).
- ^ Drumheller, Stephanie K.; McHugh, Julia B.; Kane, Miriam; Riedel, Anja; D’Amore, Domenic C. High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem. PLOS ONE. May 27, 2020, 15 (5): e0233115. Bibcode:2020PLoSO..1533115D. ISSN 1932-6203. PMC 7252595 . PMID 32459808. doi:10.1371/journal.pone.0233115 (英语).
- ^ Norman, D.B. Carnosaurs. The Illustrated Encyclopedia of Dinosaurs. Salamander Books Ltd. 1985: 62–67. ISBN 978-0-517-46890-6.
- ^ Senter, P. Vestigial skeletal structures in dinosaurs. Journal of Zoology. 2010, 280 (4): 60–71. doi:10.1111/j.1469-7998.2009.00640.x.
- ^ Sanders, R.K.; Smith, D.K. The endocranium of the theropod dinosaur Ceratosaurus studied with computer tomography. Acta Palaeontologica Polonica. 2005, 50 (3).
- ^ 57.0 57.1 57.2 Molnar, R.E. Theropod paleopathology: a literature survey. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 337–363.
- ^ Rothschild, B.; Tanke, D.H.; Ford, T.L. Theropod stress fractures and tendon avulsions as a clue to activity. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 331–336.
- ^ Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations. Geological Society of America Abstracts with Programs. 2006, 38 (6): 7.
- ^ Bilbey, S.A. Cleveland-Lloyd Dinosaur Quarry – age, stratigraphy and depositional environments. Carpenter, K.; Chure, D.; Kirkland, J.I. (编). The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. 1998: 87–120. ISSN 0026-7775.
- ^ Russell, D.A. An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. 1989: 64–70. ISBN 978-1-55971-038-1.
- ^ 62.0 62.1 Mateus, O. Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 223–231.
- ^ Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- ^ Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. The fauna and flora of the Morrison Formation: 2006. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 233–248.
- ^ Carpenter, K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 131–138.
- ^ Mateus, O.; Mannion, P.D.; Upchurch, P. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology. 2014, 34 (3): 618–634. doi:10.1080/02724634.2013.822875.
- ^ A Smithsonian Dino-Celebrity Finally Tells All. [2022-07-10]. (原始内容存档于2022-07-13).
- ^ Lucas, F. A. (1901). Animals of the Past. McClure, Phillips.
- ^ Hutchinson, H. N. (1910). Extinct Monsters and Creatures of Other Days: A Popular Account of Some of the Larger Forms of Ancient Animal Life. Chapman & Hall.
- ^ The Gilmore Models: Where are they now?. [2022-07-10]. (原始内容存档于2022-07-14).
- ^ Title: The Land That Time Forgot. isfdb.org. [2022-08-01]. (原始内容存档于2022-11-20).
- ^ Giovanni Drogo (D. Buzzati), The killing of the dragon , in Oggi, Rizzoli, 3 June 1939, p. 8.
- ^ Dupuy, Georges. Newnes, George , 编. The Monster of 'Partridge Creek'. The Strand Magazine. Vol. 36 no. 211 (George Newnes Ltd.). July 1908: 73–79 –通过Internet Archive.
- ^ Dupuy, Georges. Le Monstre de 'Partridge Creek' [The Monster of "Partridge Creek"]. Je sais tout. Vol. 4 no. 39 (Paris, France: Pierre Lafitte & Cie). 15 April 1908: 403–409 [2022-08-01]. (原始内容存档于2022-09-03) –通过Bibliothèque nationale de France (法语).
- ^ Lydekker, Richard. The Monster of Partridge Creek. Zoology. Knowledge & Illustrated Scientific News (London, England). September 1908, 5 (9): 219 –通过Hathi Trust.
- ^ Primitive Man, (a.k.a. Brute Force) at silentera.com. [2022-08-01]. (原始内容存档于2020-10-28).
- ^ Ceratosaurus | Jurassic Park Wiki - Fandom. [2022-08-01]. (原始内容存档于2022-10-19).
- ^ The Animal World (页面存档备份,存于互联网档案馆)
- ^ When Dinosaurs Roamed America (页面存档备份,存于互联网档案馆)